by Ali Attar
Nominated by Daisy Haas for CHEM 353: Introduction to Biochemical Research Techniques and Scientific Writing
Instructor Introduction
Ali's review article on 'M2 Protein: The Henchman of Influenza A' highlights his consideration and cleverness for integrating narrative into a complex scientific review assignment. Potentially even more challenging than finding and understanding biochemical research on protein interactions is communicating that knowledge to new audiences through writing. This essay stood out, as Ali described and analyzed complex chemical structure, function, and impact on human health while maintaining approachable and creative language. Essays like these challenge the field of science writing to create more connections with the public, striving to break down barriers in communication between scientists and the general public. I am honored to nominate Ali's review article for a Sweetland Upper-Level Writing Prize.
- Daisy Haas
M2 Protein: The Henchman of Influenza A
Abstract
Critical to the Influenza A virus is one of the smallest ion channels known, the M2 protein. The majority of the action is contained in a hydrophobic pocket between residues 22 and 27 with the histidine 37 and tryptophan 41 cluster. This pocket is pH dependent where at high pH the transmembrane domain is compact, with the histidine acting like a filter and the tryptophan a gate. At low pH the histidine gets deprotonated when a hydronium ion attacks, destabilizing the transmembrane helix and allowing the tryptophans to rotate to a parallel conformation that allows proton flow. This channel activation then allows viral replication, which can be inhibited by stopping proton conduction. By discovering drugs that inhibit the conductance of the M2 channel, we can combat the Influenza A virus.
Introduction
Achoo! As the cold months approach us leading into winter, a silent and deadly foe awaits. The flu season draws near and with it comes the countless deaths that harm people all around the world. The culprit? Influenza A, a sometimes deadly virus responsible for the majority of flu illnesses1. One of three types of influenza, type A is the most common and has the quickest change of genetic makeup as compared to the other types, making it harder to treat and more likely to induce a pandemic1. Influenza A also accounts for avian flu and other infections that can spread to animals, making it an even greater threat. As a result, common treatments include either antivirals or allowing the symptoms to go away. More research is needed to develop these medications because, without them, Influenza A is deadly and can ruin lives1.
Critical to Influenza A are the acid-activated proton channels (M2) in the viral envelopes essential for viral replication. M2 is a transmembrane protein located in the lipid bilayer of the viral envelope with differing ideas on how it works2. Sharma et. al proposes a possible mechanism for how M2 activation works through a Histidine-Tryptophan cluster believed to be responsible for the conductance mechanism. This is relevant because the “Conductance Domain” has been responsive to the antiviral drug Amantadine in the past, making M2 mutations an important focus, and there is a need for improved drugs to treat amantadine-resistant strains of the virus.
Understanding the Influenza A viral replication is critical3. This progresses through four steps: entry, activation, conductance, and uncoating. It begins with a virion entering a cell through receptor-mediated endocytosis. Viral particles enter the environment of the lumen and M2 becomes activated, allowing protons to conduct across the viral membrane which lowers the virion pH. Finally, that low pH initiates virus uncoating and releases ribonucleotides and viral machinery to enter the cell nucleus and start transcription3.
Delving deeper into the structure and function of the M2 protein, we can understand mutations that lead to disease and learn about potential targets for medications targeting the virus. Understanding aspects of the conductance domain and how pH activates the channel will deliver an understanding of key aspects of the mechanisms and will deliver future directions for the fate of combating Influenza A. For example, by inhibiting conductance, viral replication can be stopped4. As one of the smallest ion channels, M2 allows us to realize how the structure and function are related in order to develop these medications5.
The paper will begin by creating an image of the main area of focus, the hydrophobic pocket, and describing its different parts. I will then discuss the specific actions that can be taken in that area in order to deliver function for the protein. These are heavily influenced by pH and protons are of utmost importance. Specific stability and structural components will be observed in order to confer an understanding of the way our lives can be ruined by the M2 protein in Influenza A.
Discussion
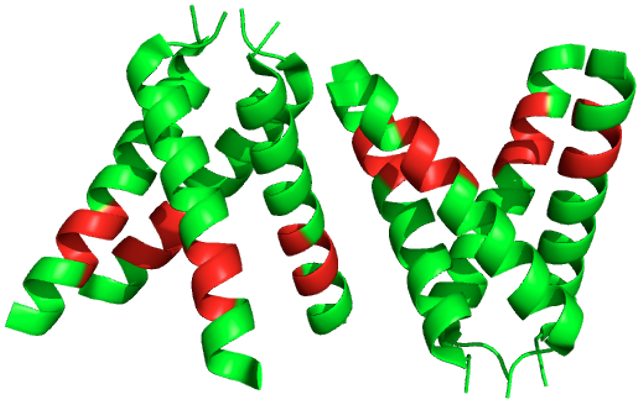
The hydrophobic pocket (shown in Figure 1) holds the majority of the action. An area once thought of as where rimantadine (an antiviral drug that treats Influenza A) binds, the hydrophobic pocket is in the transmembrane helix, which contains an aspartic acid hidden at the bottom (Figure 2)4. After researchers determined the structure, they noticed that the helices in the transmembrane tilt push the hydrophobic pocket to be wider and remove access from the aspartic acid. This suggests, for example, that rimantadine can't bind from the exterior, although it had been previously observed as being bound due to a detergent environment that had been used in a research setting to characterize the structure. This shows the importance of the environment where we look at our structures2.
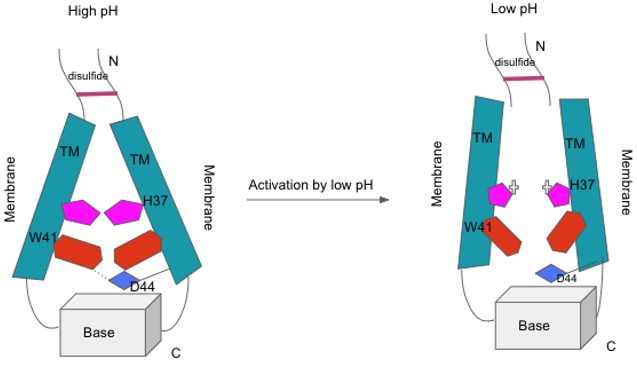
M2 channel activation. When the pH is high, the transmembrane (TM) helices are in positions that are compact and contributed to by the tryptophan gate (in red) and its association and interaction with aspartic acid (blue). At low pH, the histidine (pink) gets protonated and this destabilized the packing of the transmembrane helix, but not completely due to the N-terminal disulfide bond (top) and the base at the C-terminal (bottom) that keeps basic stability. The destability allows proton conductance to be possible.
Specifically, within the hydrophobic pocket, the HxxxW quartet is where all the action including acid activation and proton conductance happens. The HxxxW quartet is made up of the his37-trp41 cluster (shown in Figure 3). The his37 residues had pKa values around 8 and lower, meaning that the acid was relatively strong. At a pH of 7.5, the histidine was doubly protonated and shared between nearby histidines which strengthened the hydrogen bond through resonance2.
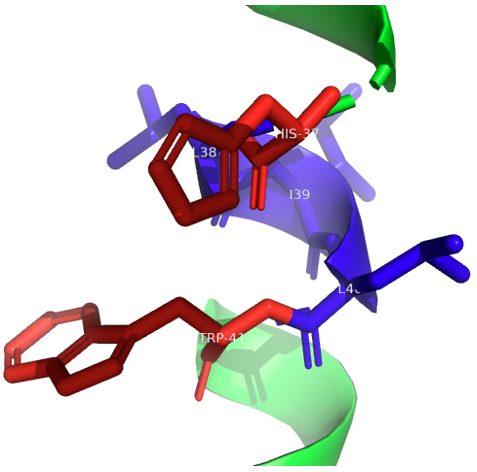
This histidine tetrad is a pair of imidazole-imidazolium dimers where the proton is shared. Here, however, it was discovered that the two imidazole rings in the backbone were parallel to each other which is not as favorable as it being perpendicular. In the dimers, multiple important atoms exist. The N sigma 1 and N epsilon 2 atoms in the dimers aren't part of the hydrogen bonds and are instead protonated and toward the C-terminal side2. The N epsilon 2 protons interact with the indole in the trp41 (the other end of the quartet), while the N sigma 1 protons make hydrogen bonds with the oxygens of the carbonyls in the histidine (in the backbone)2. To emphasize, the histidine is essentially locked in the HxxxW quartet and the channel is blocked because the protons can't be released from the imidazole N-H protons to the C-terminal pore (Figure 4).
The other imidazole nitrogens that can give protons are part of strong hydrogen bonds6. If the protons were exchanged from the N-terminal pore, and there were some side chain rotations, the imidazole-imidazolium dimers would end up exchanging partners. However, since the NMR data consistently showed a symmetric structure on average, this partner exchange happens very quickly, and almost unnoticeably2. This means that we can basically disregard the switching of partners and consider that the structure remains consistent.
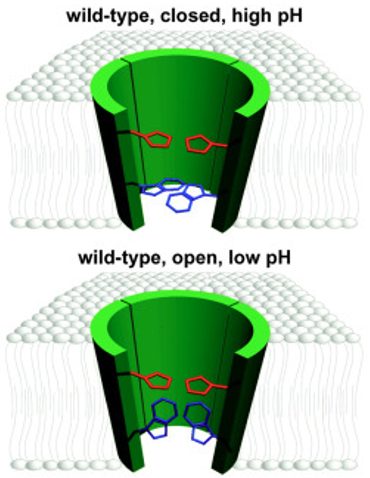
Model showing the channel structure and transmembrane domains of M2 with His37 in red and Trp41 in blue to represent the activation of M2. This shows the activation of the M2 channel and shows the transmembrane domain from 24-44 in green and His37, which acts as a filter, in red. Trp41, acts as a gate, in blue. The top depiction represents the closed channel and it has a pH on the outside where the histidine is not charged and the tryptophan is “closed” at the end of the pore. On the bottom, we see a lower pH where the histidine is charged and allows the tryptophan to rotate to a parallel conformation that allows hydrogens to flow. This is the “open” conformation6.
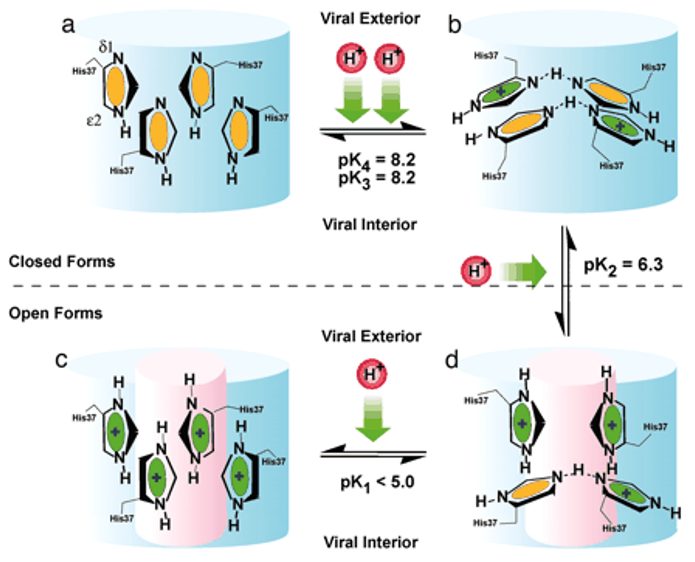
M2 channel opening. Four stages of protonation are shown: in a, all four histidines are neutral, b has two charges, c has three charges, and d has a fully protonated histidine tetrad. Neutral residues are in yellow and positively charged histidine residues are in green. The open state is the pink areas. Dashed lines in histidine dimers represent strong hydrogen bonds. This shows that when you disrupt one of the dimer structures it results in a high-energy state where the system is willing to give up a proton and return to the paired dimer state. This is supported because the second and third pKas are separated by nearly 2 pH units, so it shows how difficult it is to disrupt the stable histidine-locked conformation7.
Using a neutral pH to study the structure of the HxxxW quartet, researchers determined the mechanism of the acid activation and proton conductance. Researchers discovered that under acidic conditions in the exterior, a hydronium ion from the N-terminal pore attacks an imidazole- imidazolium dimer2. This is now the activated state and the now triply protonated histidine tetrad is stabilized by a hydrogen bond with water at the N sigma one site that there is now access to. This is located on the N-terminal side and another stabilizing factor is an additional cation pi interaction with a tryptophan 41 residue at the N epsilon 2 site on the C-terminal side2.
Using Raman spectroscopy in the transmembrane domain with acidic conditions, this strong cation pi interaction with histidine 37 and tryptophan 41 was observed2. Raman spectroscopy is a spectroscopic technique that's used to determine the vibrational modes of molecules, and rotational and other low-frequency modes of systems may also be observed. Raman spectroscopy is usually used in chemistry to provide a structural fingerprint by which molecules can be identified.
The strong cation pi interactions between the histidine 37 and tryptophan 41 protect the protons on the C-terminal side from water access2. However, when the helical backbone undergoes conformational changes, like a change in the helix kink in glycine 34, in addition to the movement of the tryptophan 41 side chain it leads to the breaking of the cation pi interaction2. This is now the conducting state and the N epsilon 2 proton is exposed to water on the C-terminal side which lets it be released to the C-terminal pore2. When that proton is released, everything repeats. Basically, the histidine becomes locked again and is ready again for proton uptake from the N-terminal, then the proton releases to the C-terminal pore. In each round of the proton conductance, as the histidine changes from being locked, activated, and conducting for the HxxxW quartet, the angle of the histidine 37 and tryptophan 41 side chains need to change and they rotate. These rotational angles, however, were discovered to be much smaller than previously thought and similarly, they are lower than what’s needed for the imidazole-imidazolium dimers when they have to exchange partners2.
So to sum it all up, acid activation in the histidine-locked state is initiated with a proton transfer from the hydronium ion into the inter-residue hydrogen bond between N sigma 1 and N epsilon 2. In the resulting activated state, the two imidazolium rings rotate so that the two nitrogens move toward the center of the pore; in addition, the protonated N sigma 1 forms a hydrogen bond with water in the N-terminal pore while the protonated N epsilon 2 moves downward (when the angle relaxes) to form a cation-pi interaction with an indole, and that blocks water access from the C-terminal pore. The conducting state is obtained when this indole moves aside to expose the N epsilon 2 proton to water in the C-terminal pore7. Once the N epsilon 2 proton is released to C-terminal water, the HxxxW quartet returns to the histidine-locked state.
Conclusion and Future Directions
Influenza A is an RNA virus that contains genetic material made up of negative sense RNA8. The M2 protein comes from the matrix protein gene that encodes different mRNAs, one of which is the M2 protein. Among many different components of the M2 protein, some portions of the structure are uniformly conserved and some aren’t. However, the HxxxW quartets are always conserved8.
Vital to the murdering nature of Influenza A is that very protein, the M2 protein, and by extension the hydrophobic pockets in the protein channels. As horrifying as having submicroscopic elements controlling our potential health may seem, knowing about the structures and functions of proton conductance allows us to understand and develop ways to combat a disease that has harrowed us for centuries. This all begins with the activation of the N-terminal due to low pH to lead to the channel conductance. Beginning as high as a pH of 8.5, conductance increases as pH drops to 4, and the protein channel is more permeable to protons than various ions9. This allows the protein to use the ions in an antiporter system to remove ions and bring in protons8.
Looking at the structure of this hydrophobic pocket, it begins at the N-terminal with a hydrophobic residue, Valine-27, and ends with the Tryptophan-41 residue. In between, from the top, the pore forms as it grows in size until Glycine-348. The HxxxW quartet is a critical function and starts with Histidine-37 which senses different protons and conducts them. In order to conduct them, the histidine uses its imidazole in its side chain which gets protonated and deprotonated. At the bottom of the pore, the tryptophan works in association with Aspartic Acid-44 to serve as a gate for the channel. In order for proton conductance, we see the histidine side chain bind a proton on the outside of the gate and release the proton that is bound on the inside.
Different drugs can mess up this process. These include Rimantadine and Amantadine8. Specifically, Amantadine can begin to harm the structure of the pocket. Typically, there is a kink at the Glycine-34 residue. This serves to prevent hydrophobic residues in the transmembrane domain from getting exposed to unfavorable environments. Amantadine can smooth out the kink normally present. In addition to its binding throughout the channel, it harms the structure by reducing the conformational flexibility of the channel8.
The M2 protein and its channel are especially important during infection. The M2 channel moves on the trans-Golgi network, a secretory pathway10. The network has a low pH. This activates the ion channel and then raises the pH of the trans-Golgi network. This then affects the cell's activity by harming the function of the secretory pathway which is also linked to homeostasis. A delay is caused in that system and we know that because by blocking the M2 channel, with a drug like Amantadine, we observed the delay removed. Following the adverse effects of making the pH of the secretory pathways higher, we see that different proton channels are affected11. One channel is the cystic fibrosis transmembrane conductance regulator, or CFTR, which is inhibited by the higher pH. It becomes inhibited due to degradation by the lysosome. The M2 protein also leads to amiloride-sensitive epithelial sodium channels, or ENaC, being affected. Reactive oxygen species are increased in the cell as the result of the pH increases which affects membrane potentials around the cell and leads to the degradation of ENaC through the stimulation of protein kinase C.
All in all, Influenza A is a killer and its main henchman is the M2 protein. As the flu season falls among us, it is out for more victims and we need more ways to combat this villain in society. By understanding the structure and function of the M2 protein and what goes on inside the channel that leads to terrible things in our cells, we can develop new ways to combat the virus and develop new drugs to ensure safety and health in our society, from the Influenza A virus.
References
(1) What to Know About Type A Influenza Symptoms, Treatment and Prevention. Healthline. https://www.healthline.com/health/influenza-a-symptoms (accessed 2022-09-26).
(2) Sharma, M.; Yi, M.; Dong, H.; Qin, H.; Peterson, E.; Busath, D. D.; Zhou, H.-X.; Cross, T. A. Insight into the Mechanism of the Influenza A Proton Channel from a Structure in a Lipid Bilayer. Science 2010, 330 (6003), 509–512. https://doi.org/10.1126/science.1191750.
(3) Takeda, M.; Pekosz, A.; Shuck, K.; Pinto, L. H.; Lamb, R. A. Influenza A Virus M 2 Ion Channel Activity Is Essential for Efficient Replication in Tissue Culture. J Virol 2002, 76 (3), 1391–1399. https://doi.org/10.1128/JVI.76.3.1391-1399.2002.
(4) Schnell, J. R.; Chou, J. J. Structure and Mechanism of the M2 Proton Channel of Influenza A Virus. Nature 2008, 451 (7178), 591–595. https://doi.org/10.1038/nature06531.
(5) Cady, S. D.; Luo, W.; Hu, F.; Hong, M. Structure and Function of the Influenza A M2 Proton Channel. Biochemistry 2009, 48 (31), 7356–7364. https://doi.org/10.1021/bi9008837.
(6) Tang, Y.; Zaitseva, F.; Lamb, R. A.; Pinto, L. H. The Gate of the Influenza Virus M2 Proton Channel Is Formed by a Single Tryptophan Residue. Journal of Biological Chemistry 2002, 277 (42), 39880–39886. https://doi.org/10.1074/jbc.M206582200.
(7) Hu, J.; Fu, R.; Nishimura, K.; Zhang, L.; Zhou, H.-X.; Busath, D. D.; Vijayvergiya, V.; Cross, T. A. Histidines, Heart of the Hydrogen Ion Channel from Influenza A Virus: Toward an Understanding of Conductance and Proton Selectivity. Proc. Natl. Acad. Sci. U.S.A. 2006, 103 (18), 6865–6870. https://doi.org/10.1073/pnas.0601944103.
(8) Manzoor, R.; Igarashi, M.; Takada, A. Influenza A Virus M2 Protein: Roles from Ingress to Egress. IJMS 2017, 18 (12), 2649. https://doi.org/10.3390/ijms18122649.
(9) Chizhmakov, I. V.; Geraghty, F. M.; Ogden, D. C.; Hayhurst, A.; Antoniou, M.; Hay, A. J. Selective Proton Permeability and PH Regulation of the Influenza Virus M2 Channel Expressed in Mouse Erythroleukaemia Cells. The Journal of Physiology 1996, 494 (2), 329–336. https://doi.org/10.1113/jphysiol.1996.sp021495.
(10) T, S.; Gp, L.; Ra, L. The Ion Channel Activity of the Influenza Virus M2 Protein Affects Transport through the Golgi Apparatus. The Journal of cell biology 1996, 133 (4). https://doi.org/10.1083/jcb.133.4.733.
(11) Londino, J. D.; Lazrak, A.; Jurkuvenaite, A.; Collawn, J. F.; Noah, J. W.; Matalon, S. Influenza Matrix Protein 2 Alters CFTR Expression and Function through Its Ion Channel Activity. American Journal of Physiology-Lung Cellular and Molecular Physiology 2013, 304 (9), L582–L592. https://doi.org/10.1152/ajplung.00314.2012.